Human leukocyte antigen

The human leukocyte antigen system (HLA) is the name of the major histocompatibility complex (MHC) in humans. The superlocus contains a large number of genes related to immune system function in humans. This group of genes reside on chromosome 6, and encode cell-surface antigen-presenting proteins and many other genes. The HLA genes are the human versions of the MHC genes that are found in most vertebrates (and thus are the most studied of the MHC genes). The proteins encoded by certain genes are also known as antigens, as a result of their historic discovery as factors in organ transplantations. The major HLA antigens are essential elements for immune function. Different classes have different functions:
HLA class I antigens (A, B & C) present peptides from inside the cell (including viral peptides if present). These peptides are produced from digested proteins that are broken down in the proteasomes. The peptides are generally small polymers, about 9 amino acids in length. Foreign antigens attract killer T-cells (also called CD8 positive- or cytotoxic T-cells) that destroy cells.
HLA class II antigens (DP,DM, DOA,DOB,DQ, & DR) present antigens from outside of the cell to T-lymphocytes. These particular antigens stimulate T-helper cells to multiply, and these T-helper cells then stimulate antibody-producing B-cells to produce Antibodies to that specific antigen. Self-antigens are suppressed by suppressor T-cells.
HLA class III antigens encode components of the complement system.
HLA have other roles. They are important in disease defense. They may be the cause of organ transplant rejections. They may protect against or fail to protect (if down regulated by an infection) cancers[1]. They may mediate autoimmune disease (examples: type I diabetes, coeliac disease). Also, in reproduction, HLA may be related to the individual smell of people and may be involved in mate selection [2].
Aside from the genes encoding the 6 major antigens, there are a large number of other genes, many involved in immune function, located on the HLA complex. Diversity of HLA in human population is one aspect of disease defense, and, as a result, the chance of two unrelated individuals having identical HLA molecules on all loci is very low. Historically, HLA genes were identified as a result of the ability to successfully transplant organs between HLA similar individuals.
Contents |
Functions
The proteins encoded by HLAs are those on the outer part of body cells that are (effectively) unique to that person. The immune system uses the HLAs to differentiate self cells and non-self cells. Any cell displaying that person's HLA type belongs to that person (and therefore is not an invader).

In infectious disease. When a foreign pathogen enters the body, specific cells called antigen-presenting cells (APCs) engulf the pathogen through a process called phagocytosis. Proteins from the pathogen are digested into small pieces (peptides) and loaded onto HLA antigens (specifically MHC class II). They are then displayed by the antigen presenting cells for certain cells of the immune system called T cells, which then produce a variety of effects to eliminate the pathogen.
Through a similar process, proteins (both native and foreign, such as the proteins of viruses) produced inside most cells are displayed on HLA antigens (specifically MHC class I) on the cell surface. Infected cells can be recognized and destroyed by components of the immune system (specifically CD8+ T cells).
The image off to the side shows a piece of a poisonous bacterial protein (SEI peptide) bound within the binding cleft portion of the HLA-DR1 molecule. In the illustration far below, a different view, one can see an entire DQ with a bound peptide in a similar cleft, as viewed from the side. Disease-related peptides fit into these 'slots' much like a hand fits into a glove or a key fits into a lock. In these configurations peptides are presented to T-cells. The T-cells are restricted by the HLA molecules when certain peptides are within the binding cleft. These cells have receptors that are like antibodies and each cell only recognizes a few class II-peptide combinations. Once a T-cell recognizes a peptide within an MHC class II molecule it can stimulate B-cells that also recognize the same molecule in their sIgM antibodies. Therefore these T-cells help B-cells make antibodies to proteins they both recognize. There are billions of different T-cells in each person that can be made to recognize antigens, many are removed because they recognize self antigens. Each HLA can bind many peptides, and each person has 3 HLA types and can have 4 isoforms of DP, 4 isoforms of DQ and 4 Isoforms of DR (2 of DRB1, and 2 of DRB3,DRB4, or DRB5) for a total of 12 isoforms. In such heterozygotes it is difficult for disease related proteins to escape detection.
In graft rejection. Any cell displaying some other HLA type is "non-self" and is an invader, resulting in the rejection of the tissue bearing those cells. Because of the importance of HLA in transplantation, the HLA loci are among of the most frequently typed by serology or PCR relative to any other autosomal alleles.
| HLA allele | Diseases with increased risk | Relative risk |
|---|---|---|
| HLA-B27 | Ankylosing spondylitis | 12 |
| Postgonococcal arthritis | 14 | |
| Acute anterior uveitis | 15 | |
| HLA-DR3 | Autoimmune hepatitis | 14 |
| Primary Sjögren syndrome | 10 | |
| Diabetes mellitus type 1 | 5 | |
| HLA-DR4 | Rheumatoid arthritis | 4 |
| Diabetes mellitus type 1 | 6 | |
| HLA-DR3 and-DR4 combined | Diabetes mellitus type 1 | 15 |
| HLA-B47 | 21-hydroxylase deficiency | 15 |
| Unless else specified in boxes, then ref is: [3] | ||
In autoimmunity. HLA types are inherited, and some of them are connected with autoimmune disorders and other diseases. People with certain HLA antigens are more likely to develop certain autoimmune diseases, such as Type I Diabetes, Ankylosing spondylitis, Celiac Disease, SLE (Systemic Lupus Erythematosus), Myasthenia Gravis, inclusion body myositis and Sjögren's syndrome. HLA typing has led to some improvement and acceleration in the diagnosis of Celiac Disease and Type 1 diabetes; however for DQ2 typing to be useful it requires either high resolution B1*typing (resolving *0201 from *0202), DQA1*typing, or DR serotyping. Current serotyping can resolve, in one step, DQ8. HLA typing in autoimmunity is being increasingly used as a tool in diagnosis. In Celiac disease it is the only effective means of discriminating between 1st degree relatives who are at risk from those who are not at risk, prior to the appearance of sometimes irreversible symptoms such as allergies and secondary autoimmune disease.
In cancer. Some HLA mediated diseases are directly involved in the promotion of cancer. Gluten sensitive enteropathy is associated with increased prevalence of Enteropathy-associated T-cell Lymphoma, and DR3-DQ2 homozygotes are within the highest risk group with close to 80% of gluten sensitive EATL cases. More often, however, HLA molecules play a protective role, recognizing the increase in antigens that were not tolerated because of low levels in the normal state. Abnormal cells may be targeted for apoptosis mediating many cancers before clinical diagnosis. Prevention of cancer may be a portion of heterozygous selection acting on HLA.
Classification
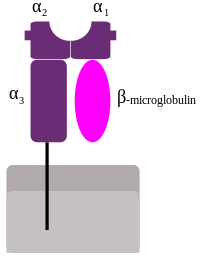
MHC class I proteins form a functional receptor on most nucleated cells of the body.
There are 3 major and 3 minor MHC class I genes in HLA:
- HLA-A
- HLA-B
- HLA-C
- minor genes are HLA-E, HLA-F and HLA-G
- β2-microglobulin binds with major and minor gene subunits to produce a heterodimer

There are 3 major and 2 minor MHC class II proteins encoded by the HLA. The genes of the class II combine to form heterodimeric (αβ) protein receptors that are typically expressed on the surface of antigen presenting cells.
Major MHC class II
- HLA-DP
- α-chain encoded by HLA-DPA1 locus
- β-chain encoded by HLA-DPB1 locus
- HLA-DQ
- α-chain encoded by HLA-DQA1 locus
- β-chain encoded by HLA-DQB1 locus
- HLA-DR
- α-chain encoded by HLA-DRA locus
- 4 β-chains (only 3 possible per person), encoded by HLA-DRB1, DRB3, DRB4, DRB5 loci
The other MHC class II proteins, DM and DO, are used in the internal processing of antigens, loading the antigenic peptides generated from pathogens onto the HLA molecules of antigen-presenting cell.
Nomenclature
Modern HLA alleles are typically noted with a variety of levels of detail. Most designations begin with HLA- and the locus name, then * and some (even) number of digits specifying the allele. The first two digits specify a group of alleles. Older typing methodologies often could not completely distinguish alleles and so stopped at this level. The third through fourth digits specify a synonymous allele. Digits five through six denote any synonymous mutations within the coding frame of the gene. The seventh and eighth digits distinguish mutations outside the coding region. Letters such as L, N, Q, or S may follow an allele's designation to specify an expression level or other non-genomic data known about it. Thus, a completely described allele may be up to 9 digits long, not including the HLA- prefix and locus notation.
Variability

MHC loci are some of the most genetically variable coding loci in mammals, and the human HLA loci are no exceptions. Despite the fact that the human population went through a constriction more than 150 000 years ago that was capable of fixing many loci, the HLA loci appear to have survived such a constriction with a great deal of variation.[4] Of the 9 loci mentioned above, most retained a dozen or more allele-groups for each locus, far more preserved variation than the vast majority of human loci. This is consistent with a heterozygous or balancing selection coefficient for these loci. In addition, some HLA loci are among the fastest evolving coding regions in the human genome. One mechanism of diversification has been noted in the study of Amazonian tribes of South America that appear to have undergone intense gene conversion between variable alleles and loci within each HLA gene class.[5] Less frequently, longer range productive recombinations through HLA genes have been noted producing chimeric genes.
Five loci have over 100 alleles that have been detected in the human population, of these the most variable are HLA B and HLA DRB1. As of 2004, the number of alleles that have been determined are listed in the table below. To interpret this table, it is necessary to consider that an allele is a variant of the nucleotide (DNA) sequence at a locus, such that each allele differs from all other alleles in at least one (single nucleotide polymorphism, SNP) position. Most of these changes result in a change in the amino acid sequences that result in slight to major functional differences in the protein.
There are issues that limit this variation. Certain alleles like DQA1*0501 and DQA1*0505 encode proteins with identically processed products. Other alleles like DQB1*0201 and DQB1*0202 produce proteins that are functionally similar. For class II (DR, DP and DQ), amino acid variants within the receptor's peptide binding cleft tend to produce molecules with different binding capability.
Tables of variant alleles
Number of variant alleles at class I loci according to the IMGT-HLA database, last updated January 2009:
| MHC class I | |
|---|---|
| locus | #[6][7] |
| Major Antigens | |
| HLA A | 767 |
| HLA B | 1,178 |
| HLA C | 439 |
| Minor Antigens | |
| HLA E | 9 |
| HLA F | 21 |
| HLA G | 43 |
Number of variant alleles at class II loci (DM, DO, DP, DQ, and DR):
| MHC class II | ||||
|---|---|---|---|---|
| HLA | -A1 | -B1 | -B3 to -B51 | Potential |
| locus | #[7] | #[7] | #[7] | Combinations |
| DM- | 4 | 7 | 28 | |
| DO- | 12 | 9 | 72 | |
| DP- | 27 | 133 | 3,591 | |
| DQ- | 34 | 96 | 3,264 | |
| DR- | 3 | 618 | 82 | 2,121 |
| 1DRB3, DRB4, DRB5 have variable presence in humans | ||||
Sequence Feature Variant Type (SFVT)
The large extent of variability in HLA genes poses significant challenges in investigating the role of HLA genetic variations in diseases. Typically disease association studies treat each HLA allele as a single complete unit, which does not illuminate the parts of the molecule associated with disease. Karp D. R. et al. describes a novel Sequence Feature Variant Type (SFVT) approach for HLA genetic analysis which categorizes HLA proteins into biologically relevant smaller sequence features (SFs), and their variant types (VTs). [8] Sequence features are combinations of amino acid sites defined based on structural information (e.g. beta-sheet 1), functional information (e.g. peptide antigen binding), and polymorphism. These sequence features can be overlapping and continuous or discontinuous in the linear sequence. Variant types for each sequence feature are defined based upon all known polymorphisms in the HLA locus being described. SFVT categorization of HLA is applied in genetic association analysis so that the effects and roles of the epitopes shared by several HLA alleles can be identified. Sequence features and their variant types have been described for all classical HLA proteins; the international repository of HLA SFVTs will be maintained at IMGT/HLA database. [9] A tool to convert HLA alleles into their component SFVTs can be found on the Immunology Database and Analysis Portal (ImmPort) website. [10]
Examining HLA types
Serotype and allele names
There are two parallel systems of nomenclature that are applied to HLA. The, first, and oldest system is based on serological (antibody based) recognition. In this system antigens were eventually assigned letters and numbers (e.g. HLA-B27 or, shortened, B27). A parallel system was developed that allowed more refined definition of alleles, in this system a "HLA" is used in conjunction with a letter * and four or more digit number (e.g. HLA-B*0801, A*68011, A*240201N N=Null) to designate a specific allele at a given HLA locus. HLA loci can be further classified into MHC class I and MHC class II (or rarely, D locus). Every two years a nomenclature is put forth to aid researchers in interpreting serotypes to alleles.[6]
Serotyping
In order to create a typing reagent, blood from animals or humans would be taken, the blood cells allowed to separate from the serum, and the serum diluted to its optimal sensitivity and used to type cells from other individuals or animals. Thus serotyping became a way of crudely identifying HLA receptors and receptor isoforms. Over the years serotyping antibodies became more refined as techniques for increasing sensitivity improved and new serotyping antibodies continue to appear. One of the goals of serotype analysis is to fill gaps in the analysis. It is possible to predict based on 'square root','maximum-likelihood' method, or analysis of familial haplotypes to account for adequately typed alleles. These studies using serotyping techniques frequently revealed, particularly for non-European or north East Asian populations a large number of null or blank serotypes. This was particularly problematic for the Cw locus until recently, and almost half of the Cw serotypes went untyped in the 1991 survey of the human population.
There are several types of serotypes. A broad antigen serotype is a crude measure of identity of cells. For example HLA A9 serotype recognizes cells of A23 and A24 bearing individuals, it may also recognize cells that A23 and A24 miss because of small variations. A23 and A24 are split antigens, but antibodies specific to either are typically used more often than antibodies to broad antigens.
Cellular typing
| HLA-DR | Cellular specificity |
|---|---|
| DR1 | Dw1, Dw20 |
| DR2 | Dw2, Dw12, Dw21, Dw22 |
| DR3 | Dw3 |
| DR4 | Dw4, Dw10, Dw13, Dw14, Dw15 |
| DR11(5) | Dw5 |
| DR13(6) | Dw, Dw18(w6), Dw19(w6) |
| DR14(6) | Dw9, Dw16 |
| DR7 | Dw7, Dw11(w7), Dw17(w7) |
| DR8 | Dw8 |
| DR9 | Dw23 |
| DR52 | Dw24, Dw25, Dw26 |
A representative cellular assay is the mixed lymphocyte culture (MLC) and used to determine the HLA class II types.[11] The cellular assay is more sensitive in detecting HLA differences than serotyping. This is because minor differences unrecognized by alloantisera can stimulate T cells. This typing is designated as Dw types. Serotyped DR1 has cellularly defined as either of Dw1 or Dw20 and so on for other serotyped DRs. Table[12] shows associated cellular specificities for DR alleles. However, cellular typing has inconsistence in the reaction between cellular type individuals sometimes resulting differently from predicted. Together with difficulty of cellular assay in generating and maintaining cellular typing reagents, cellular assay is being replaced by DNA-based typing method.[11]
Gene sequencing
Minor reactions to subregions that show similarity to other types can be observed to the gene products of alleles of a serotype group. The sequence of the antigens determines the antibody reactivities and so having a good sequencing capability (or sequence based typing) obviates the need for serological reactions. Therefore different serotype reactions may indicate the need to sequence a persons HLA to determine a new gene sequence. Broad antigen types are still useful, such as typing very diverse populations with many unidentified HLA alleles (Africa, Arabia,[13] Southeastern Iran[14] and Pakistan, India[15]). Africa, Southern Iran and Arabia shows the difficulty in typing areas that were settled earlier, allelic diversity makes it necessary to use broad antigen typing followed by gene sequencing because there is an increased risk of misidentifying by serotyping techniques.
In the end, a workshop, based on sequence, decides which new allele goes into which serogroup either by sequence or reactivity. Once the sequence is verified it is assigned a number. For example, a new allele of B44 may get a serotype B*4465 as it is the 65th B44 allele discovered. Marsh et al. (2005)[6] can be considered a code book for HLA serotypes and genotypes and a new book biannually with monthly updates in Tissue Antigens.
Phenotyping
Gene typing is different from gene sequencing and serotyping. With this strategy PCR primers specific to a variant region of DNA are used (called SSP-PCR), if a product of the right size is found, the assumption is that the HLA allele has been identified. New gene sequences often result in an increasing appearance of ambiguity. Because gene typing is based on SSP-PCR it is possible that new variants, particularly in the class I and DRB1 loci may be missed.
For SSP-PCR within the clinical situation is often used for identifying HLA phenotypes. An example of an extended phenotype for a person might be:
A*0101/*0301, Cw*0701/*0702, B*0702/*0801, DRB1*0301/*1501, DQA1*0501/*0102, DQB1*0201/*0602
This is generally identical to the extended serotype: A1,A3,B7,B8,DR3,DR15(2), DQ2,DQ6(1)
For many populations such as the Japanese or European populations so many patients have been typed that new alleles are relatively rare, and thus SSP-PCR is more than adequate for allele resolution. Haplotypes can be obtained by typing family members. In areas of the world where SSP-PCR is unable to recognize alleles and typing requires the sequencing of new alleles. Areas of the world were SSP-PCR or serotyping may be inadequate include Central Africa, Eastern Africa, parts of southern Africa, Arabia and S. Iran, Pakistan and India.
Haplotypes
An HLA haplotype is a series of HLA "genes" (loci-alleles) by chromosome, one passed from the mother and one from the father.
The phenotype exampled above is one of the more common in Ireland and is the result of two common genetic haplotypes:
A*0101 : Cw*0701 : B*0801 : DRB1*0301 : DQA1*0501 : DQB1*0201 (By serotyping A1-Cw7-B8-DR3-DQ2)
which is called ' 'super B8' ' or ' 'ancestral haplotype' ' and
A*0301 : Cw*0702 : B*0702 : DRB1*1501 : DQA1*0102 : DQB1*0602 (By serotyping A3-Cw7-B7-DR15-DQ6 or the older version "A3-B7-DR2-DQ1")
These haplotypes can be used to trace migrations in the human population because they are often much like a fingerprint of an event that has occurred in evolution. The Super-B8 haplotype is enriched in the Western Irish, declines along gradients away from that region and is only found in areas of the world where Western Europeans have migrated. The "A3-B7-DR2-DQ1" is more widely spread, from Eastern Asia to Iberia. The Super-B8 haplotype is associated with a number of diet associated autoimmune diseases. There are 100000s of extended haplotypes but only a few show a visible and nodal character in the human population.
Role of allelic variation
Studies of humans and other animals imply a heterozygous selection mechanism operating on these loci as an explanation for this exceptional variability.[16] One credible mechanism is sexual selection in which females are able to detect males with different HLA relative to their own type.[17] While the DQ and DP encoding loci have fewer alleles combinations of A1:B1 can produce a theoretical potential of 1586 DQ and 2552 DP αβ heterodimers, respectively. While certainly nowhere near this number of isoforms exist in the human population, each individual can carry 4 variable DQ and DP isoforms increasing the potential number of antigens that these receptors can present to the immune system in individual immune system. Studies of the variable positions of DP, DR, and DQ reveal that peptide antigen contact residues on class II molecules are most frequently the site of variation in the protein primary structure. Therefore, through a combination of intense allelic variation and/or subunit pairing the class II 'peptide' receptors are capable of binding an almost endless variation of peptides of 9 amino acids or longer in length, protecting interbreeding subpopulations from nascent or epidemic diseases. Individuals in a population frequently have different haplotypes, and this results in many combinations, even in small groups. This diversity enhances the survival of such groups, and thwarts evolution of epitopes in pathogens, which would otherwise be able to be shielded from the immune system.
Antibodies
HLA antibodies are typically not naturally occurring, with few exceptions are formed as a result of an immunologic challenge of a foreign material containing non-self HLAs via blood transfusion, pregnancy (paternally-inherited antigens), or organ or tissue transplant.
Antibodies against disease associated HLA haplotypes have been proposed as a treatment for severe autoimmune diseases.[18]
Donor-specific HLA antibodies have been found to be associated with graft failure in kidney, heart, lung and liver transplantation.
HLA matching for sick siblings
In some diseases requiring hematopoietic stem cell transplantation, preimplantation genetic diagnosis may be used to give rise to a sibling with matching HLA, although there are ethical considerations. [19]
Further reading
- Harsanyi, Zsolt; Hutton, Richard (1982). Genetic Prophecy: Beyond the Double Helix. London: Granada. ISBN 0-246-11760-5
External links
- IMGT/HLA Sequence Database at European Bioinformatics Institute
- hla.alleles.org
- HLA Informatics Group at The Anthony Nolan Trust
- American Society for Histocompatibility and Immunogenetics
- European Federation for Immunogenetics
- HistoCheck HLA matching tool for organ and stem cell transplantation
- Allele Frequencies at Variable Immune related loci
- MeSH Human+leukocyte+antigens
References
- ↑ Bottley G, Watherston O, Hiew Y, Norrild B, Cook G and Blair G (2007). "High-risk human papillomavirus E7 expression reduces cell-surface MHC class I molecules and increases susceptibility to natural killer cells". Oncogene. [Epub ahead of print] PMID 1782829.
- ↑ Brennan P and Kendrick K (2006). "Mammalian social odours: attraction and individual recognition". Philos Trans R Soc Lond B Biol Sci. 361(1476):2061-78 PMID 17118924
- ↑ Table 5-7 in: Mitchell, Richard Sheppard; Kumar, Vinay; Abbas, Abul K.; Fausto, Nelson (2007). Robbins Basic Pathology. Philadelphia: Saunders. ISBN 1-4160-2973-7. 8th edition.
- ↑ Shennan, Douglas H (2006). Evolution and the Spiral of Technology. Trafford Publishing. ISBN 1552125181.
- ↑ P. Parham and T. Ohta (1996). "Population Biology of Antigen Presentation by MHC class I Molecules". Science 272 (5258 pages = 67–74): 67. doi:10.1126/science.272.5258.67. PMID 8600539..
- ↑ 6.0 6.1 6.2 Marsh SG, Albert ED, Bodmer WF, Bontrop RE, Dupont B, Erlich HA, Geraghty DE, Hansen JA, Hurley CK, Mach B, Mayr WR, Parham P, Petersdorf EW, Sasazuki T, Schreuder GM, Strominger JL, Svejgaard A, Terasaki PI, and Trowsdale J. (2005). "Nomenclature for factors of the HLA System, 2004". Tissue Antigens 65 (4): 301–369. doi:10.1111/j.1399-0039.2005.00379.x. PMID 15787720.
- ↑ 7.0 7.1 7.2 7.3 IMGT/HLA Database
- ↑ Karp DR, Marthandan N, Marsh SG, Ahn C, Arnett FC, Deluca DS, Diehl AD, Dunivin R, Eilbeck K, Feolo M, Guidry PA, Helmberg W, Lewis S, Mayes MD, Mungall C, Natale DA, Peters B, Petersdorf E, Reveille JD, Smith B, Thomson G, Waller MJ, and Scheuermann RH. (2010). "Novel sequence feature variant type analysis of the HLA genetic association in systemic sclerosis. 2010". Human Molecular Genetics 19 (4): 707–719. doi:10.1093/hmg/ddp521. PMID 19933168.
- ↑ "IMGT/HLA Database". http://www.ebi.ac.uk/imgt/hla/.
- ↑ "Immunology Database and Analysis Portal (ImmPort)". https://www.immport.org/immportWeb/home/home.do?loginType=full.
- ↑ 11.0 11.1 Hurley CK (1997). "DNA-based typing of HLA for transplantation." In Leffell MS, Donnenberg AD, Rose NR, eds. (1997) Handbook of Human Immunology. pp. 521-55, Boca Raton: CRC Press, ISBN 0849301343.
- ↑ Bodmer JG, Marsh SG, Albert ED, Bodmer WF, Dupont B, Erlich HA, Mach B, Mayr WR, Parham P, Sasazuki T, Schreuder GMT, Strominger JL, Svejgaard A, Terasaki PI (1992). "Nomenclature for factors of the HLA system, 1991." Human Immunol 34(1): 4-18, PMID 1399721.
- ↑ Valluri V, Mustafa M, Santhosh A, Middleton D, Alvares M, El Haj E, Gumama O, and Abdel-Wareth L (2005). "Frequencies of HLA-A, HLA-B, HLA-DR, and HLA-DQ phenotypes in the United Arab Emirates population". Tissue Antigens 66 (2): 107–113. doi:10.1111/j.1399-0039.2005.00441.x. PMID 16029430.
- ↑ Farjadian S, Naruse T, Kawata H, Ghaderi A, Bahram S, and Inoko H (2004). "Molecular analysis of HLA allele frequencies and haplotypes in Baloch of Iran compared with related populations of Pakistan". Tissue Antigens 64 (5): 581–587. doi:10.1111/j.1399-0039.2004.00302.x. PMID 15496201.
- ↑ Shankarkumar U, Prasanavar D, Ghosh K, and Mohanty D (2003). "HLA A*02 allele frequencies and B haplotype associations in Western Indians". Hum Immunol. 64 (5): 562–566. doi:10.1016/S0198-8859(03)00032-6. PMID 12691707.
- ↑ V. Apanius, D. Penn, P.R. Slev, L.R. Ruff, and W.K. Potts (1997). "The nature of selection on the major histocompatibility complex". Critical Reviews in Immunology 17 (2): 179–224. PMID 9094452..
- ↑ Wedekind C, Seebeck T, Bettens F, and Paepke AJ (1995). "MHC-dependent mate preferences in humans". Proc Biol Sci. 260 (1359): 245–249. doi:10.1098/rspb.1995.0087. PMID 7630893.
- ↑ Oshima M, Deitiker P, Ashizawa T, Atassi M (2002). "Vaccination with a MHC class II peptide attenuates cellular and humoral responses against tAChR and suppresses clinical EAMG". Autoimmunity 35 (3): 183–90. doi:10.1080/08916930290022270. PMID 12389643.
- ↑ Verlinsky Y, Rechitsky S, Schoolcraft W, Strom C, Kuliev A (Jun 2001). "Preimplantation diagnosis for Fanconi anemia combined with HLA matching". JAMA 285 (24): 3130–3. doi:10.1001/jama.285.24.3130. PMID 11427142. http://jama.ama-assn.org/cgi/pmidlookup?view=long&pmid=11427142.
|
||||||||||||||||||||||